
 詹妮弗·邦纳
詹妮弗·邦纳
生物学副教授
地址:
生物系- CIS 210C
火博体育大学
萨拉托加泉,纽约州12866
电话:
(518) 580-5089
传真:
(518) 580-5071
个人网站:
http://jbonner.weebly.com/
电子邮件:
jbonner@mulher-perfeita.net
教育及专业培训:
新罕布什尔大学,微生物学学士
英属哥伦比亚大学神经科学博士
美国犹他大学博士后
课程:
生物科学2
247细胞生物学
【火博体育官网】神经发育
分子神经科学的342前沿
385分子生物学与遗传学、综合生物学和生态学方法
进化与行为
神经科学101神经科学入门
神经科学综合研讨会
主要研究方向:
我研究的长期目标是确定控制神经系统的基因 连接. 我用斑马鱼作为模型系统来解决这个问题 重点研究脊髓,因其结构相对简单,具有较高的同源性 脊椎动物包括人类. 几个进化上保守的神经元类别 都存在于斑马鱼的脊髓中,包括联合神经元,感觉神经元, 和运动. 联合神经元连接着神经系统的两个部分 并通过对吸引和排斥信号的反应来发展,这允许他们 越过动物的中间,称为中线. 感觉神经元前突 在脊髓后部,同时也延伸到皮肤上,形成了广泛的结构 乔木. 最后,运动神经元细胞体,位于脊髓腹侧,突出 从脊髓到肌肉组织. 我对引导线索很感兴趣 指导这些神经元亚型的寻路. 为了做到这一点,我使用了遗传学, 基因敲低技术与细胞生物学研究神经元寻路机制 探索经典指导线索的潜在新角色.
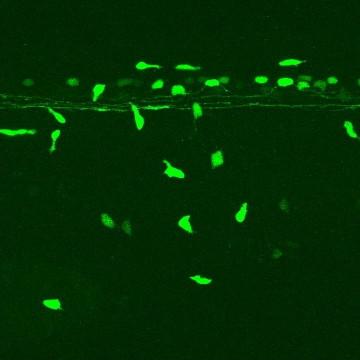
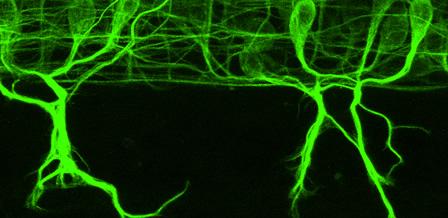
GFP标记脊髓神经元后向投射时的共聚焦时间序列 在斑马鱼胚胎中. 两条脊髓束从右侧(前)进入野区。 脊髓的一侧. 大的GFP标记的外胚层细胞也可以看到迁移 整个胚胎. (点击图片观看视频)
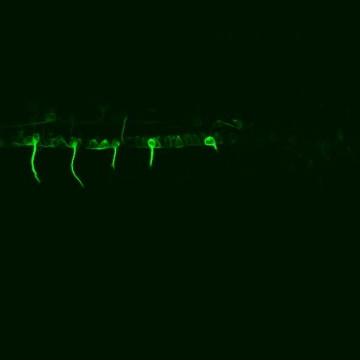
gfp标记的斑马鱼运动神经元支配周围环境的延时共聚焦电影 肌肉组织. (点击图片观看视频)
同行评审出版物:
贝克A,瓦特,R. 邦纳J. (2014)安装斑马鱼胚胎以获得最佳显微镜. 杂志 视觉实验(84):50703.
罗斯, 邦纳J. (2012)使用氯化锂激活Wnt信号:一个基于查询的模型 本科生实验练习. 斑马鱼,准备好了.
斯普纳点, 邦纳J, Maricq AV, Benian GM, Norman KR. (2012)需要UNC-89-obscurin的大型同工异构体 对秀丽隐杆线虫肌肉细胞结构和钙释放的影响. 公共科学图书馆7 (7):e40182
邦纳J, Letko米Nikolaus OB; 克鲁格L, 库珀一, 查德威克B, 康克林P,林爱华,陈国斌,杜国荣. (2012)后续不需要中线交叉 联合神经元的寻路决策. 神经发育7(1):18
格里布尔SL,金HS, 邦纳J王晓明,王晓明. (2009) Tcf3通过调节sox4a抑制脊髓神经发生 表达式. 发展136 (5):781 - 9.
邦纳J, Gribble SL, Veien ESNikolaus OB; Weidinger G, Dorsky RI. (2008)扩散 图案是在脊髓背侧下游独立调节的 典型Wnt信号. 生物工程学报,31 (1):398- 397.
刘易斯杰, 邦纳J, Modrell M, Ragland JW, Moon RT, Dorsky RI, Raible DW. (2004)重申Wnt 斑马鱼神经嵴发育过程中的信号传导. 发展131 (6),1299 - 1308.
邦纳J, Gerrow KA和O'Connor TP. (2003) 在活的有机体内 神经元生长和引导模型. 细胞生物学71:171-193.
邦纳J,奥尔德VJ和奥康纳TP. (2002)迁移的中胚层形成均匀分布 发育中的蚱蜢胚胎中的层粘连蛋白. 生物工程学报,2009 (1):57-73.
邦纳J 和奥康纳TP. (2001)准许的线索层粘连蛋白是生长锥转动所必需的 在活的有机体内. 神经科学学报,21(24):982 - 991.
邦纳J 和奥康纳TP. (2000)信号蛋白在无脊椎动物外周发育中的作用 神经系统. 生物化学学报[j] (5): 663 - 668.
霍纳王伟,法尔凯,杨晨,王涛,谢军, 邦纳J ,胡丽,林志强,吕志强,林志强. (1997)扣带皮层突触末端 精神分裂症的蛋白质和神经细胞粘附分子. 神经科学学报[j] (1): 99-110.
最近的实验室学生











